[Anàlisi de les Dades]
[La corba de vol i l'índex d'abundància]
La metodologia del BMS s’ha dissenyat per estimar quantitativament i de manera senzilla els canvis numèrics en les poblacions de ropalòcers. Amb aquest objectiu, al final de cada temporada es calcula, per a cada espècie i estació, un índex anual d'abundància que inclou els valors estimats per a les setmanes en què no es disposa de comptatges.
L'estimació d' aquests comptatges no existents s'aconsegueix amb màxima fiabilitat a partir de la modelització de la corba de vol de les espècies mitjançant el mètode denominat GAM regional (Schmucki et al., 2016). Aquest mètode consisteix en ajustar, cada temporada, un model general additiu als comptatges setmanals d’una espècie als diferents itineraris que pertanyen a una regió climàtica concreta. D'aquesta manera, es calcula una corba fenològica única per regió i temporada que, adaptada segons l’abundància de l’espècie a cada localitat, permet estimar els comptatges de les setmanes no mostrejades.
La suma dels mostreigs reals i estimats és el que proporciona l'índex anual d'abundància d'una espècie en una localitat. Aquests càlculs es duen a terme amb el paquet estadístic rbms (Schmucki et al., 2021).
[Les regions climàtiques]
Tant la fenologia com les dinàmiques poblacionals de les papallones es veuen molt influïdes pel clima. Per aquest motiu, les dades s'analitzen a nivell de tres regions climàtiques, en cadascuna de les quals s’aplica un model GAM per determinar la corba fenològica regional de l'espècie en qüestió.
La regió climàtica a la qual pertany un itinerari s’ha establert segons un criteri tèrmic. Concretament el número d'hores per any en què se supera una temperatura de 21ºC (21DDG; segons dades proporcionades pel Servei Meteorològic de Catalunya).
D'aquesta manera, s'han diferenciat tres regions climàtiques a partir dels següents llindars: regió alpina i subalpina (≤ 0-150 DDG), regió mediterrània humida (entre 150-400 DDG), regió mediterrània àrida (≥ 400 DDG). Aquestes regions es corresponen amb força exactitud a les regions bioclimàtiques definides per Metzguer et al. (2013), àmpliament utilitzades en estudis de modelització ecològica. Com s'ha comentat més amunt, es considera que les poblacions d'una espècie tenen la mateixa corba de vol en el conjunt d'estacions del CBMS d'una certa regió climàtica.
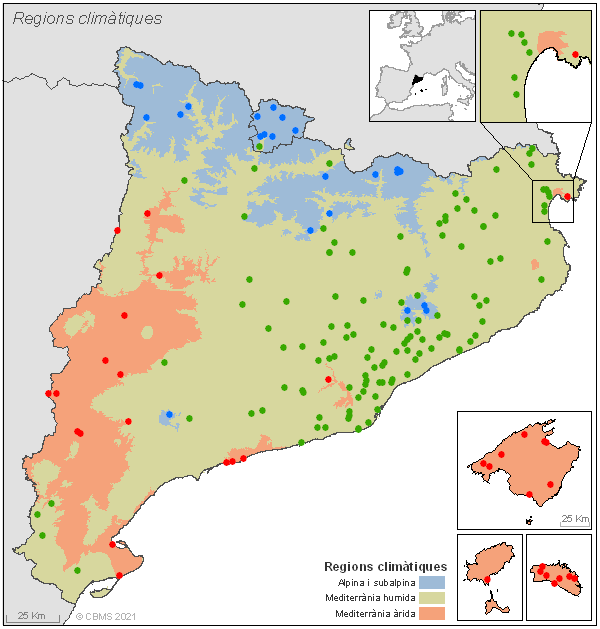
Mapa de la xarxa CBMS, mostrant les estacions de seguiment fins a 2020 i les tres regions climàtiques a les quals pertanyen segons el model climàtic de Metzguer et al. (2013).
[La corba de vol]
Per a estimar la corba de vol regional d'una espècie cada temporada, s’utilitzen les dades d'aquells itineraris on es considera que l'espècie hi manté poblacions viables. Hem adoptat com a criteri de població viable que almenys hi hagi dues ocurrències de l'espècie en una temporada de mostreig. Així mateix, la corba de vol només es calcula en el cas que hi hagi un mínim de cinc itineraris amb poblacions viables en almenys un dels anys de mostreig.
Encara que les corbes de vol d'una espècie per regió s'estimen anualment, a les fitxes d’espècie es mostra una corba de vol mitjana que sintetitza el patró fenològic observat pel conjunt d'anys disponibles.
La corba de vol de la blanqueta perfumada (Pieris napi) s'ha pogut estimar per a dues regions climàtiques, l’alpina i la mediterrània humida. La fenologia difereix en aquestes dues regions. Mentre que a la regió mediterrània humida la corba de vol presenta tres pics ben diferenciats i una màxima abundància al mes de juny, a la regió alpina la corba de vol és més tardana i presenta només dos pics i un màxim al mes d'agost-setembre. La interpretació de les corbes fenològiques pot ser complexa ja que no sempre un pic es correspon inequívocament amb una generació. Per exemple, en aquesta figura, el màxim poblacional del juny-juliol a la regió mediterrània humida podria correspondre alguns anys a dues generacions àmpliament superposades.
[Les tendències poblacionals]
[Tendències locals]
A nivell de cada itinerari, s'obtenen els índexs d’abundància que informen de l'abundància relativa anual d'una espècie en aquella localitat. Aquests índexs es calculen a partir de la suma dels comptatges a l'itinerari. En aquells casos en què ha estat possible estimar la corba de vol regional mitjançant el model GAM, els índexs anuals inclouen els comptatges reals i els imputats per a les setmanes perdudes. En cas contrari, només inclouen els comptatges reals. D'altra banda, si les dades reals corresponen a un percentatge molt baix (menor al 10%), es considera que el model no és prou fiable per a fer un càlcul d’un índex d’abundància (l'espècie, en aquest cas, apareix com a no avaluada).
A les fitxes d'espècie per itinerari es mostren els índexs anuals estandarditzats a una distància fixa de 100 m de transsecte per facilitar la comparació entre poblacions de diferents itineraris. Els valors d'abundància d'una espècie que apareixen a la taula general a les fitxes d'espècie i a les fitxes d'itinerari corresponen a mitjanes per a tots els anys disponibles.
Al CBMS es calcula un únic índex anual d'abundància per espècie, que reflecteix l'èxit de l'espècie al llarg de tot el període de vol comprès dins de les 30 setmanes oficials de la temporada. Si la fenologia és suficientment clara com per detectar generacions discretes, els índexs anuals d'abundància es podrien calcular per separat per a cada generació. No obstant, donada la complexitat que presenten les fenologies als diferents climes del territori català, es descarta aquesta opció per als càlculs actuals. Existeix, a més a més, una certa variabilitat segons les condicions meteorològiques de la temporada i són freqüents les espècies bivoltines amb una tercera generació parcial, així com les polivoltines amb un nombre variable de generacions. En altres projectes de BMS, com l’UKBMS al Regne Unit, sí que es distingeixen índexs anuals per generació, i en les espècies bivoltines s'utilitza únicament l'índex de la segona generació, generalment més abundant.
Els índexs d'abundància local s'utilitzen per calcular les tendències locals de les espècies als diferents itineraris. Aquestes tendències s'estimen a partir d’una regressió lineal entre el logaritme de l'índex local (log [IA+1]) i l'any, sempre que la sèrie temporal disposi de vuit o més anys durant els quals l'espècie hagi aparegut com a mínim quatre anys seguits. En cas contrari, l'espècie apareix com a "No avaluada". A les diferents fitxes, les tendències locals apareixen classificades segons el pendent de la recta de regressió i el seu nivell de significació en les categories:
- Regressió forta: declivi significatiu i amb més d’un 20% de canvi en un període de 20 anys.
- Regressió moderada: declivi significatiu i amb menys d’un 20% de canvi en un període de 20 anys.
- Increment moderat: augment significatiu i amb menys d’un 20% de canvi en un període de 20 anys.
- Increment fort: augment significatiu i amb més d’un 20% de canvi en un període de 20 anys.
- Estable: tendència no significativa amb intervals de confiança petits que permeten afirmar que hi hauria menys d’un 20% de canvi en un període de 20 anys.
- Incerta: tendència no significativa i intervals de confiança prou grans com per considerar que la tendència podria suposar un canvi major que el 20% en un període de 20 anys.
- Extingida: quan s’ha enregistrat un fenòmen d’extinció local sense una posterior recolonització. El criteri utilitzat per definir una extinció és la seqüència de quatre anys seguits sense l'aparició d’una espècie en una localitat, precedits per una seqüència de quatre anys amb presència d’aquella espècie en aquell indret.
Els gràfics de tendències inclouen sèries temporals no actuals quan una estació ja fa temps que no és activa. En tals casos, la tendència fa referència al període de mostreig disponible en aquella estació, sense indicar en quin estat es troba l’espècie actualment.
Tendències poblacionals de dues espècies de satirins a l'itinerari del Cortalet, PN Aiguamolls de l'Empordà. La saltabardisses europea, Pyronia tithonus, va patir una forta davallada la segona meitat de la dècada dels 2000, que va acabar comportant l'extinció de la població local. Contràriament, la bruixa, Brintesia circe, va colonitzar la zona l'any 2004, i en els darrers anys ha estat capaç d'establir una població creixent. En ambdós casos, les tendències semblen ser una resposta als factors climàtics, ja que els hàbitats on es presenten les dues espècies han patit poques alteracions en aquest transsecte.
[Tendències regionals]
La xarxa del BMS recull dades relatives a un nombre elevat d'estacions i, per tant, cal disposar d'un mètode que permeti el tractament simultani de tota aquesta informació. Amb aquesta finalitat, es calcula un índex anual regional que agrupa les dades de totes les estacions d'una regió climàtica i que s'utilitza per conèixer la tendència d'una espècie al llarg del temps en aquesta regió. Per a estimar un índex regional s’ha determinat el mínim de cinc itineraris amb dades de poblacions viables, de la mateixa manera que amb les corbes de vol.
Per calcular l'índex anual regional d'una espècie i la seva tendència temporal s'utilitza el paquet estadístic rbms (Schmucki et al., 2021). Per col·lapsar en un sol valor les dades de tots els itineraris on l'espècie apareix, l'índex regional anual s'estima a partir d'un model lineal generalitzat (GLM). Aquest model corregeix el biaix que comporta que alguns itineraris disposin de més dades que d’altres, repartint el pes de les dades locals segons el percentatge de la corba mostrejada en cada localitat. Alhora, el model també estima indirectament el valor que correspondria als anys en què no s'arriba al mínim de cinc itineraris pel càlcul de la corba de vol. Els índexs regionals anuals es presenten a més amb interval de confiança del 95%, calculats aquests amb un mètode bootstrap que repeteix fins a 500 vegades els càlculs amb submostres aleatòries de les dades.
Les tendències poblacionals regionals es mostren només quan es disposa d'un mínim de quatre anys seguits amb dades calculades o estimades segons els següents criteris:
- El primer any de la sèrie de l'índex regional s'ha de calcular amb un mínim de cinc itineraris.
- Els anys posteriors també han de superar aquest mínim o, en cas contrari, no hi pot haver més de dos anys seguits amb dades insuficients.
- Quan les sèries temporals queden interrompudes més de dos anys, el primer any de la sèrie passa a ser el que assegura que a partir d'aquell punt no hi haurà discontinuïtats més grans de dos anys.
- En el cas de sèries temporals incompletes (en les quals falten dades d’alguns anys), les tendències poblacionals es calculen sempre i quan hi hagi més anys calculats que anys estimats.
Les tendències poblacionals es calculen per separat per a les tres regions climàtiques definides (vegeu l'apartat Les regions climàtiques) però també per al conjunt de totes les estacions estudiades.
La safranera pàl·lida (Colias alfacariensis) es troba actualment en regressió moderada al conjunt de la xarxa CBMS. A nivell de regió climàtica, en canvi, s'observa una regressió moderada a la regió mediterrània humida però no a la regió alpina, on la tendència (disponible per a un nombre d'anys més curt) resta incerta. Cliqueu les categories de la llegenda per activar/desactivar les sèries.
[Preferències i indicadors d'hàbitat]
El CBMS recull informació precisa de l'abundància de les espècies en diferents tipus d'hàbitat, ja que els comptatges es diferencien a nivell de secció dins d'un itinerari. Aquestes dades permeten calcular diversos índexs que detallen les preferències d'una espècie per a cada ambient i que, després, formen la base per al càlcul dels indicadors multiespecífics d'hàbitat.
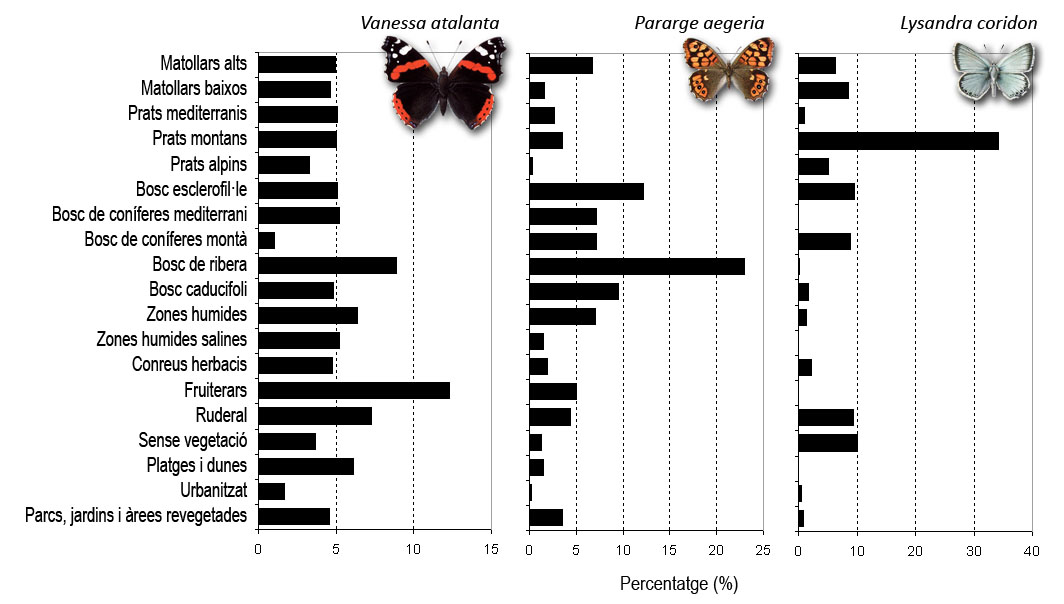
La figura mostra tres espècies ben diferents pel que fa les seves preferències d’hàbitat. L’atalanta, Vanessa atalanta, és una espècie generalista que trobem indistintament a gairebé a qualsevol tipus d’ambient per la seva gran capacitat dispersiva. La bruna de bosc, Pararge aegeria, és una papallona que apareix sobretot en la majoria d’hàbitats de tipus forestal, on és molt ben representada. En canvi, la griseta de muntanya, Lysandra coridon, és una espècie especialista dels prats montans de Pirineu i Prepirineu
[L'índex d'especialització d'hàbitat (SSI)]
Els comptatges distingeixen entre seccions que estan dominades per comunitats vegetals que es poden assignar a grans tipus d'hàbitats. Això permet explorar quins són els ambients on les poblacions d'una determinada espècie assoleixen les màximes densitats. La caracterització de les comunitats vegetals de les seccions dels itineraris (que es fa segons el codi CORINE), es va iniciar l'any 2000 i es revisa a cada itinerari cada 6 anys. Aquesta informació és essencial per després calcular indicadors d’hàbitat i detectar problemàtiques que afectin el conjunt de la biodiversitat en determinats tipus d’ambients.
Llegiu-ne més...
Les comunitats vegetals originals classificades segons el codi CORINE s’han agrupat en 19 categories d'hàbitat més àmplies per a establir una classificació útil per a l’estudi dels ropalòcers. Aquestes són:
- Matollars alts
- Matollars baixos
- Prats mediterranis
- Prats montans
- Prats alpins
- Boscos esclerofil·les
- Boscos de coníferes mediterranis
- Boscos de coníferes montans
- Boscos de ribera
- Boscos caducifolis
- Zones humides
- Zones humides salines
- Zones humides de muntanya
- Conreus herbacis
- Fruiterars
- Ruderal
- Sense vegetació
- Platges i dunes
- Urbanitzat
- Parcs, jardins i àrees revegetades
A l'apartat resultats per hàbitat trobareu una descripció detallada de cadascuna de les categories.
La preferència d'una espècie per un hàbitat es calcula a partir de la densitat que presenta en aquest hàbitat al conjunt de la xarxa del CBMS, comparada amb les densitats a la resta dels hàbitats. A efectes gràfics, les densitats per hàbitats es mostren com un percentatge respecte la suma total de densitats als 20 hàbitats definits.
El procés de càlcul és el següent. Primerament s’ha obtingut la densitat mitjana (mitjana anual del nombre d’exemplars en 100 m de transsecte) als diferents hàbitats que hi ha en un itinerari concret. Aquesta densitat s'ha calculat a partir de les dades per secció, multiplicant la mitjana de la densitat anual en aquella secció pel percentatge de recobriment dels hàbitats que hi són presents. S'ha establert un llindar de cobertura mínima del 25% d’un tipus d’hàbitat per secció, eliminant així de l’anàlisi hàbitats que són minoritaris i que molt possiblement no expliquen la presència d’una determinada espècie en aquella secció.
En una segona fase, es tenen en compte tots els itineraris on apareix l’espècie, per a fer un càlcul de la seva densitat mitjana en cadascun dels 20 hàbitats en el conjunt de la xarxa del CBMS. En aquest procés s'assigna una densitat de zero en aquells hàbitats on l’espècie no ha aparegut, de manera que finalment les densitats s'expressen en forma percentual per a un total fix de 19 hàbitats.
Aquests valors mostren la distribució de tots els individus d'una espècie detectats al CBMS entre les 19 categories d'hàbitats. Quan una espècie té una preferència molt acusada per un hàbitat concret, la majoria dels exemplars apareixen lligats a aquest hàbitat, mentre que els percentatges d'aparició a la resta dels hàbitats són molt baixos. En canvi, els percentatges de les aparicions queden molt més repartits entre les diferents categories quan una espècie és generalista. Per quantificar aquestes preferències es calcula l'índex d'especialització d'hàbitats (SSI), com el coeficient de variació (CV = desviació estàndard/mitjana) de la distribució de les densitats (expressades en percentatge) en els 20 hàbitats. Quan més especialista és una espècie, més heterogènia és la distribució de les densitats i més alt és el SSI. Contràriament, les espècies generalistes tenen densitats similars entre molts hàbitats i, per tant, el seu SSI és baix. Posteriorment, per a les fitxes de les espècies, s’han reescalat els valors per obtenir visualment un nombre categòric on assignar l’espècie. Aquesta va de l’1 (les més generalistes) al 10 (les més especialistes).
Els càlculs de preferències d'hàbitat i l'índex SSI només s'han dut a terme per a aquelles espècies que apareixen en un mínim de 5 itineraris. Així mateix, no s'han tingut en compte les dades d'una espècie en un tipus d'hàbitat determinat quan aquest hàbitat només apareix en un sol itinerari. Amb aquests llindars s'aconsegueix reduir les associacions poc realistes entre espècies de papallones i determinats hàbitats. Tot i així, cal tenir en compte que els valors de preferència d'hàbitat encara poden presentar biaixos deguts a la mida de la mostra emprada i a característiques particulars de les dades del CBMS (p. ex. concentracions en abeuradors, comportament d'encimballament, etc.), i trobar així abundàncies inusuals d'una espècie per motius aliens a l'hàbitat. La qualitat de les dades augmenta, doncs, a mesura que les sèries temporals dels itineraris són més llargues, i quantes més estacions hi ha disponibles per a una determinada espècie.
[Índex Tancat – Obert (TAO)]
L’índex TAO situa el conjunt d’espècies que apareixen a la xarxa en un gradient de preferència per hàbitats més tancats o oberts. El valor atorgat a cada espècie se situa en un rang de valors pròxims al -1 per a les espècies amb preferència d’hàbitats tancats i amb valors més pròxims a l’1 per a la preferència per hàbitats oberts (Ubach et al., 2020)..
Llegiu-ne més...
Per a calcular-lo, es realitza primer de tot una classificació binària de totes les comunitats vegetals representades a la xarxa segons si són obertes, a les quals s’assigna el valor de 1, o tancades, a les quals s’assigna el valor de -1. Seguidament, per a cada secció es calcula un valor final multiplicant els percentatges de cobertura de les comunitat vegetals presents pel valor binari que els correspon (-1 o +1). Les seccions que tenen un valor final positiu es consideren obertes, mentre que les que tenen un número final negatiu es consideren tancades. Per tal d’evitar el problema de què seccions amb valors molt pròxims al 0 es classifiquin com obertes o tancades essent un entremig de les dues situacions, hem aplicat un filtre: només s'han considerat aquelles seccions en què el valor final és ≥ |0.1|. Per tant, s'han descartat les seccions amb valors compresos en el rang (-0.1, 0.1).
Un cop feta la reclassificació de totes les seccions en obertes o tancades, per a cada itinerari s'han calculat dues mitjanes de densitats de l’espècie: una en el conjunt de les seccions obertes (Dobert), i l'altra en el de les seccions tancades (Dtancat). L'índex TAO es calcula aleshores a partir d'aquests dues mitjanes segons la fórmula desenvolupada per Suggit et al. (2012).
![]()
El valor final de l’índex TAO d'una espècie és la mitjana dels índexs TAO obtinguts en cadascun dels itineraris.
Altres consideracions que s’han tingut en compte per al càlcul inclouen el criteri del mínim de 5 itineraris en els que una espècie ha de ser present (igual que es fa en el càlcul de l’índex SSI i les preferències d’hàbitat).
A la fitxa de les espècies l’índex TAO es reescala atorgant un valor entre 1 i 10, que determina que les espècies amb valors més pròxims a l’1 són aquelles amb preferència per hàbitats més tancats i les més pròximes al 10 les que tenen preferència per hàbitats més oberts.
[Altres indicadors d'hàbitat]
La informació sobre la preferència d'hàbitat de les espècies es pot utilitzar per desenvolupar indicadors d'hàbitat, que combinen les dades poblacionals d’un grup d’espècies característiques per conèixer el comportament de la biodiversitat en relació amb un cert ambient. Alguns dels indicadors d'hàbitat que es generen amb les dades del CBMS i BMSAnd són l'indicador europeu de papallones de prats, l'indicador dels efectes del tancament forestal i l'indicador d'hàbitats del Departament de Territori i Sostenibilitat de la Generalitat de Catalunya.
[Referències recomanades]
- Julliard, R., Clavel, J., Devictor, V., Jiguet, F., Couvet, D. (2006) Spatial segregation of specialists and generalists in bird communities. Ecology Letters 9:1237–1244
- Melero, Y., Stefanescu, C., Pino, J. (2016) General declines in Mediterranean butterflies over the last two decades are modulated by species traits. Biological Conservation, 201: 336-342. DOI 10.1016/j.biocon.2016.07.029
- Metzger, M.J., Bunce, R.G.H., Jongman, R.H.G., Sayre,R., Trabucco, A., Zomer. R. (2013) A high-resolution bioclimate map of the world: a unifying framework for global biodiversity research and monitoring. Global Ecology and Biogeography, 22: 630-638.
- Schmucki, R., Pe'er, G., Roy, D.B., Stefanescu, C., Van Swaay, C.A.M., Oliver, T.H., Kuussaari, M., Van Strien, A., Ries, L., Settele, J., Musche, M., Carnicer, J., Schweiger, O., Brereton, T., Harpke, A., Heliölä, J., Kühn, E., Julliard, R (2016) Regionally informed abundance index for supporting integrative analyses across butterfly monitoring schemes. Journal of Applied Ecology, 53: 501-510. DOI 10.1111/1365-2664.12561
- SCHMUCKI R., HARROWER C.A., DENNIS E.B. (2021) rbms: Computing generalised abundance indices for butterfly monitoring count data. R package version 1.1.0. https://github.com/RetoSchmucki/rbms
- Stefanescu, C., Torre, I, Jubany, J., Páramo, F. (2011) Recent trends in butterfly populations from north-east Spain and Andorra in the light of habitat and climate change. Journal of Insect Conservation 15: 83-93. DOI 10.1007/s10841-010-9325-z
- Ubach, A., Páramo, F., Gutiérrez, C., Stefanescu, C. (2020) Vegetation encroachment drives changes in the composition of butterfly assemblages and species loss in Mediterranean ecosystems. Insect Conservation and Diversity, 13: 151-161. DOI 10.1111/icad.12397
[Web map] | [Notes legals] | Créditos
[Project in agreement with:]



En colaboración con:

